Los sistemas de memoria son esenciales para nuestra capacidad de almacenar, retener y recuperar información. Los sistemas de memoria son procesos cognitivos que permiten codificar, almacenar y recuperar información, y se clasifican en diferentes tipos según su duración y el tipo de información procesada.
La memoria es una función cerebral fascinante por medio de la cual el sistema nervioso puede codificar, almacenar, organizar y recuperar una variedad de información relevante para el tema.
El estudio formal de la memoria comenzó hace más de un siglo, proporcionando en este tiempo una cantidad considerable de información científica sobre el funcionamiento de la memoria. El conocimiento real de la memoria nos permite considerarla lejos de ser una función única, aislada o estática, sino más bien como una compleja red de sistemas de memoria que trabajan en paralelo por un objetivo común. La evolución histórica de los conceptos de memoria y sus clasificaciones han progresado simultáneamente a nuestro conocimiento de estos sistemas.
Los primeros enfoques empíricos para la descripción de la memoria fueron encontrados en la antigua Grecia por autores como Platón y Aristóteles, y se basaron en la filosofía, el pensamiento y la introspección y los métodos lógicos. Sin embargo, los primeros enfoques científicos reales aparecieron en el siglo XIX, por autores como Ebbinghaus y Lashley, quienes iniciaron el estudio experimental de la memoria en humanos y animales, respectivamente.
La contribución del grupo intelectual conocido como <<conductista>> a principios del siglo XX, también fue relevante durante este período, y fue representada por científicos como Pavlov, Watson, Skinner y Thorndike, quienes detallaron una variante del aprendizaje reconocida en nuestros días como aprendizaje asociativo. A su vez, esta variante se puede dividir en condicionamiento clásico (asocia un estímulo con una respuesta) y condicionamiento instrumental u operante (asocia un estímulo con un comportamiento específico).
Los conductistas afirmaron que solo sobre la base observacional del comportamiento es posible conocer los procesos vinculados al aprendizaje. Sin embargo, estos autores claramente cometieron un error cuando afirmaron que el conocimiento sobre los procesos que ocurren en el cerebro siempre estará lejos de la comprensión del investigador.
Más tarde, una de las evidencias más duras para el estudio de los procesos de memoria provino de los estudios clínicos de pacientes con lesiones cerebrales focales. Penfield, Scoville y Milner, durante la década del 60, documentaron los efectos de las lesiones quirúrgicas del lóbulo temporal sobre la memoria declarativa, encontrando alteraciones selectivas, como amnesia anterógrada severa y amnesia retrógrada con gradiente temporal.
Estos hallazgos fueron acompañados también por una descripción de los subsistemas de memoria que permanecen intactos, a pesar de esas lesiones del lóbulo temporal (memoria procedimental, memoria a largo plazo, etc.). Con base en estos hallazgos, el lóbulo temporal medial se describió como una estructura clave para la adquisición de información declarativa.
Paralelamente a los estudios clínicos, en los años 60 se produjo un primer intento de coordinar la investigación psicopsiquiátrica con el conocimiento y el protocolo científico de la biología, lo que permitió el surgimiento de disciplinas conocidas como neurociencia cognitiva y psicología cognitiva o psicofisiología, ambas creadas en un esfuerzo por explorar los mecanismos celulares y moleculares responsables del almacenamiento de la información de la memoria.
La lista de hallazgos significativos proporcionados por estas dos disciplinas científicas es extensa, pero entre los más importantes se encuentran probablemente los derivados del estudio de los procesos de memoria más elementales (es decir, habituación y sensibilización) en modelos animales invertebrados, como Aplysia spp. Estas investigaciones establecieron la base para los requisitos celulares básicos para el aprendizaje elemental, así como la base molecular para la memoria a corto y largo plazo.
La conjugación de estas descripciones clínicas junto con la evidencia experimental llevó a la postulación de las primeras clasificaciones de los sistemas de memoria durante los años 80. Una de estas clasificaciones dividía los procesos de memoria en dos categorías: la memoria declarativa —la que contiene información adquirida conscientemente y fácil de verbalizar o transmitir a otras personas (este tipo de memoria también se divide en memorias semánticas y episódicas)—, y la no declarativa —que incluye aquella información que no es fácil de verbalizar o cuya adquisición es inconsciente (este tipo de memoria implica información heterogénea en la que el papel de la conciencia es complejo y permanece en el poder—. e incluye la memoria procedimental, el cebado, el condicionamiento clásico y operante, así como las formas más elementales de aprendizaje, como la habituación y la sensibilización).
Conceptualizar estos sistemas de memoria de forma aislada es un error, ya que los resultados son lo suficientemente claros como para que todos ellos trabajen juntos la mayor parte del tiempo, y funcionan de forma independiente muy raramente, algunas veces cooperando o incluso funcionando de manera competitiva. Los estudios experimentales han demostrado que el papel de los sistemas de memoria es diferente cada vez, incluso cuando dos sujetos realizan exactamente la misma tarea de aprendizaje, lo que sugiere que la codificación, la motivación y el manejo inicial de la información pueden determinar si se procesa como información procedimental, espacial o semántica.
La reciente descripción de las relaciones competitivas entre los diferentes sistemas de memoria (declarativa vs. procedimental) resultó ser un hallazgo sobresaliente, aunque en la actualidad existe evidencia clínica y experimental sólida que indica que una disminución de las funciones en los sistemas procedimentales, espaciales o declarativos puede inducir la activación de otro sistema de memoria.
Los estudios iniciales en este campo sugieren que la inactivación transitoria de las estructuras dorsales estriatales (implicadas en el rendimiento de las habilidades motoras) o las del hipocampo (implicadas en el rendimiento de las habilidades espaciales) facilitan el aprendizaje en ese sistema permaneciendo activo después del desafío farmacológico. El significado real de esta competencia entre sistemas aún no está claro, aunque se ha propuesto que ambos sistemas han evolucionado por separado en respuesta a necesidades distintas, lo que a su vez podría explicar por qué eventualmente estos sistemas pueden competir por el manejo de la información. Desafortunadamente, varios estudios han demostrado que las relaciones entre los sistemas de memoria son complicadas y poco comprendidas hasta ahora.
En neuropsicología, el estudio de estos sistemas permite comprender cómo diferentes áreas y procesos cerebrales contribuyen a diversas formas de memoria y cómo los trastornos neurológicos pueden impactar estas capacidades.
Clasificación de los Sistemas de Memoria Sensorial. Este tipo de memoria incluye:
Memoria Icónica: Relacionada con la visión, almacena información visual por menos de un segundo.
Memoria Ecoica: Relacionada con el oído, retiene información auditiva durante unos pocos segundos.
Memoria a Corto Plazo (MCP)La MCP, también conocida como memoria de trabajo, permite el almacenamiento temporal de información. Su capacidad es limitada y retiene datos por un corto período, generalmente por segundos o minutos.
Es crucial para tareas como el razonamiento y la comprensión.
Memoria a Largo Plazo (MLP)La MLP almacena información durante periodos prolongados y se subdivide en:Memoria Declarativa
Memoria Episódica: Recuerdos de experiencias personales y eventos específicos.
Memoria Semántica: Conocimiento general del mundo, como hechos y conceptos.
Memoria No Declarativa
Memoria Procedimental: Habilidades motoras y hábitos, como montar en bicicleta o tocar un instrumento.
Aprendizaje Asociativo: Condicionamiento clásico y operante.
Evaluación de los Sistemas de Memoria en Neuropsicología
La evaluación neuropsicológica utiliza pruebas específicas para medir diferentes aspectos de los sistemas de memoria. Entre las pruebas comunes se encuentran la Escala de Memoria de Wechsler (WMS) y la Prueba de Aprendizaje Verbal de California (CVLT).
Estas pruebas ayudan a identificar los déficits en los diversos sistemas de memoria y a diseñar planes de tratamiento adecuados.Trastornos de los Sistemas de Memoria
Los trastornos que afectan los sistemas de memoria pueden incluir:Amnesia: Pérdida parcial o total de la memoria, que puede ser retrógrada o anterógrada.Demencia: Un deterioro progresivo de la memoria y otras funciones cognitivas, con la enfermedad de Alzheimer como la forma más común.Trastornos Neurológicos: Lesiones cerebrales traumáticas, accidentes cerebrovasculares y enfermedades neurodegenerativas pueden afectar diferentes sistemas de memoria.Intervenciones y Tratamientos
El tratamiento de los trastornos de memoria puede incluir terapias cognitivas, programas de rehabilitación y, en algunos casos, medicamentos.
Las estrategias de compensación y las intervenciones personalizadas son fundamentales para ayudar a los pacientes a manejar sus déficits de memoria y mejorar su calidad de vida.Importancia del Estudio de los Sistemas de Memoria en Neuropsicología.
Comprender los sistemas de memoria es crucial en neuropsicología para diagnosticar y tratar eficazmente los trastornos que afectan la capacidad de recordar.
Los neuropsicólogos desempeñan un papel esencial en la evaluación y tratamiento, proporcionando intervenciones que permiten a los pacientes optimizar su funcionamiento diario.
La memoria es sin duda un proceso cerebral notable pues, mediante él, el Sistema Nervioso codifica, organiza y almacena los sucesos pasados de tal forma que en ocasiones permite recordar de manera consciente eventos en el pasado distante tan vívidamente como si se experimentaran nuevamente. Estos recuerdos con frecuencia traen consigo experiencias emocionales intensas que van desde lo maravilloso hasta lo atemorizante. La experiencia presente está inexorablemente entretejida con los recuerdos, y el significado de las personas, de las cosas y de los eventos del presente depende de algún modo de la experiencia previa, es decir, de la memoria, de modo que el presente pierde su realce sin el pasado. La memoria no es una función cerebral estática, única o aislada, se comporta más bien como un conjunto de funciones cerebrales distintas pero estrechamente interrelacionadas que están orientadas hacia un mismo fin, por lo que resulta más correcto denominarla en términos de sistemas de memoria. Podría intentar definirse someramente a la memoria como un grupo de funciones cerebrales que tienen la tarea de clasificar, codificar, almacenar y recuperar una gran diversidad de tipos de información que resultan de importancia para el organismo en particular. La taxonomía de estos sistemas ha evolucionado paralelamente al conocimiento de las bases anatómicas y fisiológicas de la memoria. Debido a su gran extensión, el estudio correcto de los procesos de memoria ha sido tema de innumerables textos especializados a lo largo del tiempo, ya que se trata de un tema que puede abordarse desde muy distintos puntos de vista (molecular, celular, neuroquímico, anatómico, evolutivo, neuropsicológico, etc.) y se encuentra en constante actualización. El presente artículo no pretende constituirse como un tratado sobre los procesos que componen o generan la memoria, sino únicamente realizar una breve reseña sobre la evolución histórica del estudio de los procesos de memoria, enunciar las bases de la taxonomía actual de una forma sencilla y práctica, y destacar algunos puntos de polémica y debate actuales al respecto.
Antecedentes y evolución histórica
El estudio teórico de la memoria y el aprendizaje se inició desde los tiempos de Platón y Aristóteles. En las teorías del conocimiento de este último se hace alusión por primera vez a distintos niveles de conocimiento (conocimiento sensible vs. entendimiento), pues para Aristóteles el verdadero saber estaba más allá de la sensación y la simple experiencia, pues además implica el conocimiento acerca de la causa y motivo de los sucesos u objetos; además señaló que el fundamento del aprendizaje y la memoria son las asociaciones entre dos sucesos (como ocurre entre el rayo y el trueno); sin embargo, hasta antes del siglo XIX el estudio de la memoria sólo se limitaba a métodos más empíricos y filosóficos que científicos, es decir, para su estudio se utilizaban métodos como la lógica, la introspección, la comparación, la reflexión, Fue hasta el siglo XIX que comenzaron los primeros reportes y estudios respecto a la memoria y sus trastornos. En realidad, la noción sobre los distintos tipos o sistemas de memoria no es nueva y se puede encontrar ya en la literatura de los siglos XVIII y XIX, en escritos psicológicos que distinguen, por ejemplo, la memoria de los hábitos (habilidades mecánicas). De esta época son de destacar los siguientes autores: Hermann Ebbinghaus, quien en 1885 fue el primero en realizar estudios experimentales sobre la memoria de repetición verbal en seres humanos, utilizando silabas sin sentido, y además describe por primera vez la mejoría progresiva del rendimiento durante la adquisición de nuevas tareas o <<curva de aprendizaje>>; William James (1890), quien en su Tratado de Psicología, fue pionero al proponer la distinción entre memoria de corto y largo plazo; Sergei Korsakoff, quien describe junto con Carl Wernicke, el síndrome amnésico (acompañado de ataxia y oftalmoplejía), que actualmente lleva su nombre (Síndrome de Wernicke–Korsakoff) y además propone el estudio de los trastornos de memoria (amnesias) como un medio importante para conocer los procesos mnemónicos normales; pero quizá uno de los exponentes más importantes de esta época, y menos reconocidos en el medio científico, es el biólogo alemán Richard Semon, al que se atribuye la autoría de uno de los términos más utilizados en la bibliografía actual de la memoria: el engrama. De hecho, en sus teorías distinguió tres aspectos diferentes que componían los procesos de memoria: la engrafía, que representaría el proceso de codificación de la información, el engrama que representaría todos los cambios que ocurren en el Sistema Nervioso y que preservan los efectos de la experiencia, y la ecforia, que representaría la recuperación de información; en su teoría también propuso que para que ocurriera la ecforia de forma eficaz (recuperación) sería necesario que se reunieran nuevamente las condiciones que prevalecían en el momento de la engrafía (adquisición); además también propuso ideas novedosas sobre el beneficio de la repetición en la memoria, sin embargo, su teoría recibió muy poco apoyo en su época lo cual se ha reflejado en el desconocimiento de sus contribuciones hasta nuestros días. A pesar de todas estas contribuciones, hasta este momento a los procesos de memoria no se les reconocía un sustrato anatómico específico dentro del Sistema Nervioso. La primera asociación anatómica entre las lesiones cerebrales focales (específicamente del lóbulo temporal) y la memoria, se atribuye a Bekhterev, en 1899, quien demostró en un paciente cuyo principal problema era una alteración grave de la memoria reciente, la presencia de lo que él denominó <<reblandecimiento>> de áreas corticales específicas como el uncus, el hipocampo y áreas adyacentes de la corteza cerebral temporal.
A principios del siglo XX, la corriente científica del <<conductismo>>, con sus principales exponentes Thorndike, Pavlov, Watson y Skinner, estudiaron las características y componentes de un tipo particular de aprendizaje y memoria: el que se deriva de la asociación repetida entre un estímulo y una respuesta (condicionamiento clásico) o entre un estímulo y una conducta (condicionamiento operante). Sin duda los conocimientos aportados por la corriente del conductismo fueron de los más importantes en los inicios del siglo XX, sin embargo, merece la pena señalar las aportaciones del novelista francés Marcel Proust (1913) quien realizó descripciones teóricas trascendentes y muy novedosas acerca del fenómeno de la experiencia consciente. En su serie de novelas que reciben el nombre de <<En busca del Tiempo perdido>> se establecen las bases de lo que él denominó <<memoria involuntaria>>, que consistía en una memoria basada en la adquisición supra consciente de información, es decir, información cuya adquisición no ha sido dirigida por la atención o por la volición inteligente del sujeto, y a la cual eventualmente puede tenerse acceso mediante algún estímulo externo que la hace evidente. Estas teorías estaban sin duda adelantadas a su tiempo y conforman hasta la actualidad uno de los terrenos más fértiles de polémica en el estudio de la memoria y su relación con la conciencia.
A pesar de todas estas aportaciones científicas fue quizá Karl Lashley, en 1929,15 el que inició formalmente el estudio experimental de la memoria con su investigación llevada a cabo en ratas, encontrando que el deterioro en el desempeño en laberintos se correlacionaba más con la extensión de las lesiones cerebrales que con la localización de las mismas. Con estos resultados se sostuvo la hipótesis de la <<equipotencialidad>> de las áreas corticales la cual establecía que, con excepción de las áreas motoras y sensitivas primarias, al resto de las funciones cerebrales complejas contribuían de igual forma todas las áreas corticales. Dicha hipótesis se encuentra en oposición directa con el concepto actual sobre los distintos sistemas y sustratos anatómicos para la memoria, aunque se trató sin duda de la primera aproximación realmente científica al estudio experimental de la memoria en el laboratorio. Simultáneamente a esta propuesta se describieron cada vez más casos clínicos en los que, contrariamente a lo establecido por esta teoría, se demostraba una asociación entre las lesiones del lóbulo temporal y las alteraciones de la memoria, así como diversos trastornos de memoria asociados a secuelas de infecciones cerebrales con afectación del lóbulo temporal. En 1949, Donald Hebb publicó su ya famoso libro The organization of behavior, a neuropsychological theory,18 donde propuso por primera vez que la memoria podría estar conformada por subsistemas de almacenamiento transitorio de información dependientes de circuitos neuronales redundantes, y otros sistemas más perdurables ligados a cambios estructurales en las neuronas tanto pre– como postsinápticas, siendo ésta la primera propuesta que aseguraba que la memoria se encontraba conformada por subsistemas de memoria, además de sugerir el sustrato morfofisiológico de la memoria de largo plazo.
Sin duda, una evidencia trascendental en el estudio de la memoria provino de la observación de las alteraciones neuropsicológicas resultantes de las lesiones quirúrgicas o traumáticas del lóbulo temporal. En 1958, Wilder Penfield y Brenda Milner describieron dos casos en los que se había realizado resección de las estructuras mediales del lóbulo temporal como tratamiento para mejorar la intensidad de las crisis epilépticas, y encontraron como secuela postquirúrgica una severa alteración para el aprendizaje de material nuevo (amnesia anterógrada), sin alteración importante del rendimiento intelectual general. Estos pacientes conservaban además la atención y la memoria inmediata. Más aún, en los casos en los que la resección fue unilateral, se encontró alteración de la memoria de forma selectiva dependiendo del tipo de material recordado; esto es, para el material verbal cuando la resección fue izquierda, y para el material no verbal (visual) cuando fue derecha. Los reportes de estos casos llamaron la atención de William Scoville, quien también realizaba una cirugía de resección temporal–medial bilateral (como alternativa a la lobotomía frontal para el alivio de la psicosis grave en pacientes con esquizofrenia). Este autor obtuvo resultados postquirúrgicos en la memoria semejantes a los reportados por Penfield y Milner. A consecuencia de esto, Milner fue invitada a estudiar los casos de Scoville y logró documentar amnesia anterógrada en todos los casos en los que la cirugía incluía el hipocampo y el giro del parahipocampo; sin embargo, no encontró ninguna alteración residual de la memoria cuando la cirugía sólo se limitaba al uncus y la amígdala lo cual concordaba con reportes previos de Sawa et El estudio, la descripción detallada y el seguimiento de uno de los casos de Scoville (el famoso paciente HM), sentaron un antecedente muy importante en el estudio de la memoria, contradiciendo de forma casi definitiva la noción de la <<equipotencialidad>> cortical cerebral, además de ayudar a esclarecer la participación del lóbulo temporal en los mecanismos de la memoria explícita. Se pueden consultar descripciones y análisis neuropsicológicos más detallados de este caso en diversos artículos publicados con anterioridad.
Por otro lado, el estudio de los pacientes con amnesia también puso en evidencia que al afectarse selectivamente la memoria para hechos recientes y conservarse relativamente intacta la memoria para hechos más remotos, existía (como ya se sospechaba), una diferencia entre el sustrato anatómico de las memorias de corto y de largo plazo. Además, se documentaron disociaciones importantes en cuanto al manejo de la información en la memoria, según el contenido de lo que es recordado; es decir, que existían algunos aspectos de ella que parecían no afectarse en los pacientes amnésicos; por ejemplo, en el estudio del paciente HM se encontró que éste era capaz de retener un número de tres cifras por un período aproximado de 15 minutos si se mantenía repitiendo dicha cifra y no se distraía su atención con otro estímulo, más allá de lo cual olvidaba por completo la información e incluso el recuerdo de haber memorizado algo. Esta disociación ya había sido propuesta con anterioridad por Drachman y Arbit en 1966. En un estudio posterior se demostró que incluso se producían resultados contrarios cuando se asignaba la tarea de recordar material que no se podía verbalizar con facilidad; en otras palabras, no podía recordar material que no era capaz de repetir y reforzar verbalmente. Más tarde también se demostró que el aprendizaje de tareas o habilidades específicas (como la escritura en espejo) se mantenía normal en las lesiones témporo–mediales, lo cual hacia suponer que el aprendizaje de hábitos y tareas motoras podía adquirirse de forma independiente del lóbulo temporal.
Posteriormente, en 1968, Warrington y Weiskrantz, usando una versión de la prueba de figuras incompletas de Gollin, demostraron que el aprendizaje de tipo perceptual está preservado en los pacientes con amnesia anterógrada, una forma de aprendizaje que posiblemente tenga su origen en áreas visuales de asociación de orden superior. Basándose en estos hallazgos clínicos, comprobados además en modelos experimentales de lesiones mediales-temporales,Larry Squire propuso en 1986 una taxonomía de la memoria, en la cual la dividía en dos grandes subsistemas: <<la memoria declarativa y la no declarativa>> (cuadro 1).

Casi al mismo tiempo, Endel Tulving propuso un modelo de la memoria que comprendía no dos, sino tres sistemas que trabajaban de forma conjunta: <<la memoria procedimental, la memoria semántica y la episódica>>. Tulving propuso que sólo la primera de ellas (procedimental) podía funcionar de forma aislada, siendo ésta la más elemental y primitiva y que podía estar presente desde los organismos más simples. La memoria procedimental comprendería el conocimiento en relación a <<saber cómo hacer las cosas>>, y según el autor, sería una forma un poco más compleja de aprendizaje que la conducta instintiva debido a que no requiere de la asociación estímulo–respuesta. La memoria semántica es la que se denomina como genérica o de hechos particulares, y se asume que no involucra ninguna estructura de eventos o ubicación témporo-espacial; un ejemplo de este tipo de memoria serían los datos concretos que memorizamos como cifras, nombres de objetos, fechas, etc. La memoria episódica es aquella que hace referencia a eventos que ocurrieron en un lugar y tiempo específicos; la recuperación de este tipo de información requiere generalmente de la creación de <<imágenes mentales>> que son evocadas en el momento y cuyo recuerdo puede incluir una gran variedad de tipos de información: visual, auditiva, olfativa, somestésica, temporal, espacial, emotiva, etc. Este proceso puede llegar a recrear el evento pasado de una forma muy vívida en el presente. Tulving describió, a su vez, tres submodalidades de estos tres sistemas, así como también lo que llamó el <<nivel de conciencia>> de cada uno de ellos:
1. La adquisición, que como su nombre lo indica, es la forma en que ingresa la información según el tipo de memoria; 2. La representación de la información en el sistema de memoria; es decir, la forma en que se codifica o clasifica la información y 3. La expresión de conocimiento; es decir, la forma en la que la información se exterioriza por el individuo34 (cuadro 2).
Uno de los defectos que presenta este sistema de clasificación es que está basado exclusivamente en la naturaleza de la información almacenada, y no considera que estos sistemas pueden funcionar de forma complementaria y conjunta, siendo a veces difícil distinguirlos en la evaluación sistemática de la memoria. En una clasificación más reciente e incluyente, Tulving y Schacter definen cinco sistemas de memoria según los mecanismos cerebrales involucrados, el tipo e información que se maneja y los principios con los que operan cada uno de los sistemas, difiriendo de la propuesta de Squire en lo siguiente:
1. Esta clasificación sólo está basada en estudios neuropsicológicos realizados en seres humanos; 2. para Tulving, la memoria de corto plazo y la episódica serían de tipo explícito, mientras que las otras serían implícitas; 3. todos los sistemas de memoria expresan funciones cognitivas a excepción de la procedimental; 4. no se incluyen formas más elementales de memoria como la habituación y la sensibilización y, además, 5. Considera la memoria episódica y la semántica como dos subsistemas independientes, proponiendo que la información accede primero a lo que llamó <<sistema de representación perceptual>> (SRP) y posteriormente a la memoria semántica, la cual a su vez, alimentaría la memoria episódica (cuadro 3).
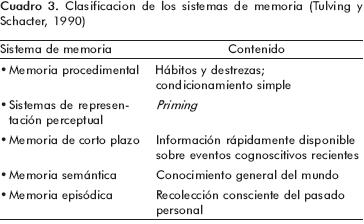
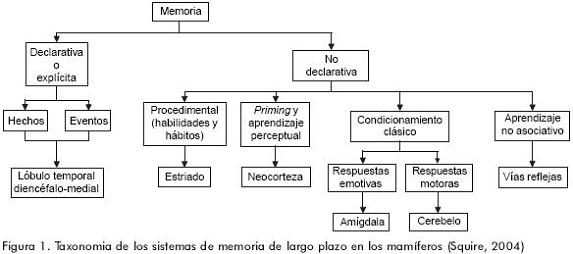
Una de las aportaciones más importantes de la clasificación de Tulving y Schacter es justamente este sistema de representación perceptual (SRP), cuyas expresiones neuropsicológicas serían la identificación perceptual y el fenómeno del priming perceptual, a cuyas características haremos referencia más adelante. En una clasificación más reciente de los sistemas de memoria de largo plazo, Larry Squire incluye además las principales estructuras anatómicas relacionadas con cada subtipo de memoria, las categorías de aprendizaje asociativo y no asociativo, así como una distinción entre los componentes emotivos y motores asociados con el condicionamiento clásico (figura 1).
El conocimiento sobre las bases moleculares de la memoria
Los estudios iniciales acerca de la memoria fueron llevados a cabo principalmente por filósofos y psicólogos, y aunque desde un inicio algunos biólogos ya habían realizado aportaciones importantes al estudio de la memoria, éstas solían recibir poco apoyo y difusión por la comunidad científica. Hasta antes de los años 1960 no existía una cooperación real y productiva entre las disciplinas biológicas y la psicología, hasta que surgió la psicología cognitiva o neurociencia cognitiva, que trataba, por primera vez, no sólo de explicar los procesos mnésicos basados en la conducta experimental observable y manipulable, sino en establecer cuáles eran los procesos celulares, y más tarde moleculares, responsables de generar y almacenar esa conducta, es decir, buscar el verdadero engrama de la memoria en el cerebro. Los fundadores de esta neurociencia cognitiva fueron George Miller, Ulric Neisser, Herbert Simons, entre otros, en la década de 1960,36 sin embargo, haciendo un análisis retrospectivo, quizá la primera propuesta acerca de los mecanismos celulares del aprendizaje la propuso Santiago Ramón y Cajal, al sugerir que el aprendizaje debe surgir del fortalecimiento en la comunicación entre las neuronas ya existentes, ya que éstas (como él observó) han perdido su capacidad de multiplicarse. Con el interés generado por el surgimiento de las neurociencias cognitivas, también se desarrollaron múltiples técnicas experimentales para el estudio neurobiológico de los procesos cognitivos. Por ejemplo, en 1970 Evarts y Mountcastle desarrollaron una técnica para el registro de la actividad eléctrica de neuronas únicas en animales vivos y en movimiento estas técnicas y las desarrolladas más tarde por Robert Wurtz, Apostolos Georgopoulos y William Newsome, produjeron los primeros estudios que correlacionaban la actividad neuronal con procesos cognitivos como la percepción, la atención y la toma de decisiones. Hubo también un resurgimiento de los estudios anatómicos correlativos, con nuevas técnicas de visualización de tractos nerviosos, y el posterior desarrollo de las técnicas de resonancia magnética y tomografía por emisión de positrones, con las cuales se complementó el estudio anatómico de los procesos cognitivos en pacientes tanto enfermos como sanos e intactos. Sin embargo, uno de los estudios más importantes sobre los mecanismos moleculares de la memoria provino de investigaciones relacionadas con las formas más elementales de la memoria no declarativa: los procesos de habituación, deshabituación y sensibilización; éstos se estudiaron en animales invertebrados con un Sistema Nervioso muy simple, conformado sólo por entre 20 000 a 100 000 células. El ejemplo más representativo es la Aplysia, aunque también se usaron otros organismos como la Hermissenda, la Pleurobranchea, la Limax e incluso la Drosophila. Las aportaciones más destacadas de estos estudios pueden resumirse en la demostración de los ensambles celulares mínimos requeridos para el aprendizaje más elemental, así como los mecanismos moleculares y neuroquímicos responsables de la memoria de corto y largo plazo. Es decir, se demostró por primera vez que los cambios estructurales responsables de la memoria de corto plazo suelen ser modificaciones covalentes de proteínas ya existentes producidas por una u otra cinasa de segundos mensajeros; mientras que la memoria de largo plazo implica la síntesis de nuevas proteínas y que el <<switch molecular>> que media el cambio de memoria de corto a largo plazo está representado por segundos mensajeros como el cAMP, proteínas señalizadoras como la MAPK o PKA, y factores de trascripción génica como el CREB. Sin embargo, los estudios sobre los mecanismos moleculares responsables de la memoria continúan en marcha, y recientemente se ha demostrado que puede existir almacenamiento de información a largo plazo a pesar de una extensa inhibición en la síntesis de proteínas. Revisar los avances recientes en el terreno de los mecanismos moleculares de la memoria trasciende a los objetivos del presente artículo, pero se pueden encontrar en la bibliografía diversas actualizaciones del tema.
TAXONOMÍA Y TÉRMINOS DE MEMORIA
Desde luego, la evolución y transformación histórica de los sistemas de clasificación no ha terminado aún, los sistemas de memoria hasta aquí mencionados forman parte de los más aceptados y difundidos en su utilización hasta el momento actual; sin embargo, los psicólogos utilizan una gran diversidad de términos para referirse a distintas variedades, características o niveles de procesos mnésicos, por lo que en la literatura especializada podemos encontrar conceptos que pueden llegar a hacer confusa la conceptualización integral de la memoria y sus distintos componentes. Entre estos conceptos hay diversas dicotomías, como la mencionada respecto a la memoria declarativa y la no declarativa, pero existen muchas otras: procesos de memoria consciente vs. inconsciente; memoria de corto vs. largo plazo; Memoria implícita vs. explícita; memoria retrospectiva vs. prospectiva; recuerdo voluntario vs. involuntario; aprendizaje intencional vs. incidental; así como diversos términos que hacen alusión a la memoria de corto plazo (de trabajo, activa) o a diversas variedades de memoria explícita de largo plazo: semántica, episódica, autobiográfica colectiva (para una revisión extensa véase: Roediger, 2008). Las definiciones y alcances de cada uno de estos términos en ocasiones son imprecisas y con frecuencia no son completamente excluyentes entre sí, pero aún así dejan ver que el estudio de los procesos mnésicos puede hacerse desde muy distintos puntos de vista.
Hasta este momento, incluso la división inicial de los sistemas de memoria (declarativa y no declarativa) continúa siendo materia de debate debido a las distintas variedades de memoria que se han observado disociadas en estudios tanto en humanos como en animales que llegan a compartir características de ambos tipos de memoria. Esta dicotomía inicial de los sistemas es, sin embargo, muy simplista para lo que se ha demostrado en la actualidad. Conceptualizar estos sistemas como aislados resulta un grave error pues es claro que todos los sistemas de memoria trabajan en paralelo, ocasionalmente de forma independiente, a veces cooperando e incluso también de forma competitiva. Resulta evidente que la participación de los distintos sistemas de memoria no siempre es la misma, incluso cuando dos individuos distintos realizan exactamente la misma tarea de aprendizaje, lo cual pone de manifiesto que la codificación, motivación o abordaje inicial de la información puede determinar si ésta es procesada como una información procedimental, espacial, semántica o de otro tipo. La descripción reciente de las relaciones competitivas de los sistemas de memoria (declarativa vs. procedimental) resultó un hallazgo sorprendente; pero ahora existe evidencia clara (tanto en animales de experimentación como en humanos), de que la disminución de las funciones de un sistema de memoria procedimental, espacial o declarativo, puede favorecer el desempeño de otro sistema de memoria. En estudios iniciales, la inactivación transitoria de estructuras estriatales dorsales (concernidas en el desempeño de habilidades y tareas motoras) o del hipocampo (concernido en el desempeño de tareas espaciales), facilitó el desempeño en el aprendizaje del sistema que no fue inactivado farmacológicamente. Por otro lado, en estudios de imagen funcional se ha encontrado además que la participación de las distintas estructuras cerebrales (estriado-hipocampo) puede ofrecer un comportamiento diferencial en relación con el tiempo, demostrándose que en el aprendizaje inicial de las tareas perceptuales o motoras existe un aumento de la activación del hipocampo, con poca participación del estriado, en tanto que se observa el patrón de activación inverso conforme la tarea se repite y se aprende. Más aún, se ha comprobado que existe una importante modulación emocional para la utilización predominante de algunos sistemas mnésicos. El significado real de esta competencia entre algunos sistemas aún no se conoce con precisión, pero se ha propuesto que ambos sistemas han evolucionado por separado en respuesta a distintas necesidades, lo cual explicaría por qué eventualmente pueden encontrarse compitiendo por el manejo de la información. Estos y otros estudios han demostrado que las relaciones entre los distintos procesos de memoria son complejas, y no del todo comprendidas. A continuación se dará una breve explicación sobre los principales procesos componentes de los sistemas de memoria, según las clasificaciones antes comentadas.
SISTEMAS DE MEMORIA DE LARGO PLAZO
Memoria declarativa
Memoria semántica: El ámbito de la memoria semántica es la información almacenada sobre las características y atributos que definen los conceptos, así como los procesos que permiten su recuperación de forma eficiente para su utilización en el pensamiento y el lenguaje. El conocimiento del sitio anatómico en donde se producen las representaciones semánticas se ha revolucionado con el uso de los estudios de neuroimagen funcional. Estos estudios han demostrado que la información sobre las características de objetos específicos que es necesaria para la generación de conceptos es almacenada dentro de los mismos sistemas neuronales que están activos durante la percepción de esos mismos estímulos. Esto se ha comprobado en estudios con tareas de nominación de colores, nominación de acciones, objetos con formas similares, objetos manipulables hechos por el hombre (herramientas), Pero ¿cómo es que se organiza la información sobre la forma de los objetos? los estudios de imagen han demostrado que se activan áreas específicas dentro de la corteza occípitotemporal cuando se utilizan ciertas categorías de objetos; por ejemplo, objetos vivos o animales (giro fusiforme lateral) vs. Objetos inanimados (giro fusiforme medial). Incluso estudios de registro unitario con electrodos profundos en pacientes epilépticos han mostrado que las neuronas responden más a una categoría específica que a un estímulo específico, sin embargo, como se demostró posteriormente, este esquema de organización no siempre funciona lo cual ha sugerido que las representaciones de diferentes categorías de objetos están distribuidas ampliamente y pueden traslaparse entre sí, dando como resultado una gran diversidad de categorías potenciales.
Es claro que el <<espacio neuronal cortical>> para almacenar todas las posibles categorías de objetos es muy limitado, por lo que debe existir alguna organización flexible (quizá basada en algunas características comunes) o priorizada para las categorías que resulten más importantes, pero si esto es así o no, aún es tema de debate. El surco temporal superior se ha asociado con la representación de movimiento de objetos y el área ventral de la corteza premotora izquierda con la representación de objetos que están asociados a movimiento.
Por otro lado, el área prefrontal inferior izquierda sería, según algunos investigadores, la responsable de la recuperación, mantenimiento, monitoreo y manipulación de las representaciones semánticas que están almacenadas en cualquier sitio; es decir, el equivalente a una memoria de trabajo semántica. De esta manera se han documentado asimetrías hemisféricas en el procesamiento semántico, encontrándose que el hemisferio izquierdo se centra en las palabras con un significado concreto, mientras que el derecho se asocia más con el procesamiento de palabras léxicamente ambiguas. En estudios recientes se ha demostrado que la activación de estructuras del lóbulo temporal (hipocampo, amígdala, corteza témporopolar) disminuye gradualmente conforme se recupera información semántica cada vez más antigua en su adquisición, observándose el patrón de activación inverso en los lóbulos parietales y frontales, lo cual sugiere que el lóbulo temporal tiene un papel limitado en el tiempo en la recuperación semántica. La relación entre memoria semántica y el lóbulo temporal también ha sido comprobada por estudios neuropatológicos de pacientes con demencia semántica, en los cuales se ha demostrado una atrofia significativa (59–65%) de estructuras temporales como el polo temporal y la corteza perirrinal, así como en otras áreas: corteza órbitofrontal, giro fusiforme, ganglios basales y sustancia blanca. En contraste, los estudios neuropatológicos no han demostrado asimetrías hemisféricas en los patrones de atrofia cerebral. También se ha demostrado recientemente, en estudios de resonancia magnética funcional, que en pacientes adultos con lesiones temporales izquierdas existe una verdadera reorganización de las áreas corticales involucradas en el procesamiento semántico normal, dicha reorganización depende de la categoría semántica específica de objetos (vegetales, animales, etc.) que tenía asignada el área cortical dañada y de las condiciones de las áreas corticales que permanecieron indemnes para suplir la actividad de las áreas corticales dañadas.
El estudio de la memoria semántica aún no está completo, pero los estudios de imagen funcional han revolucionado el conocimiento de estos procesos en los pacientes sanos; todavía quedan por dilucidar los mecanismos precisos de categorización semántica, así como la participación del resto de las áreas corticales y de las estructuras subcorticales que se activan durante el proceso de la categorización semántica.
Toda esta info es de carácter exclusivamente orientativa. Siempre resulta necesaria la consulta profesional por temas mas puntuales y personales. SI hoy te preguntas ¿Quién no puso la casa patas arriba buscando unas llaves, un libro o cualquier otro objeto que parece que se tragó la tierra? Te cuento breve, los seres humanos disponemos de un don innato para el olvido. Hasta la próxima.